No podem parlar de la història de la Terra sense parlar de fòssils. Fòssil és una paraula que prové del llatí,
fossile i que significa "el que s'extreu de la terra". Però exactament què és un fòssil?
Un fòssil és qualsevol resta d'ésser viu o de la seva activitat que queda conservada a les roques.
Ja en l'Antiguitat clàssica grans pensadors com Plini interpretaven correctament els fòssils quan els trobaven, però no va ser fins a segles posteriors en el que els avenços en l'estudi de fòssils va desenvolupar una disciplina científica especialitzada: la
Paleontologia o l'estudi dels éssers antics.
Però quants tipus de fòssils hi ha?
De fòssils n'hi ha molts tipus, podem trobar el propi ésser viu mineralitzat, un motlle extern o intern de l'ésser, una empremta o reflex de les parts orgàniques, l'esquelet extern o intern i el que anomenem
icnofòssils que són totes les empremtes de l'activitat d'un ésser viu: petjades, excrements mineralitzats, ous, nius, etc.


Fins hi tot, en casos molt excepcionals, podem trobar conservat l'ésser viu sencer tal com era quan estava viu, és el cas de vertebrats o plantes congelades en gel, com els mamuts o els insectes conservats en ambre, que és una reïna solidificada d'alguns arbres.
I com és produeix aquesta fossilització?
Per fossilitzar s'ha de donar un procés de
mineralització, durant el qual el sediment on s'ha registrat aquesta activitat o on s'han acumulat les restes queda transformat en roca sòlida preservant aquesta activitat. Així mateix la mineralització és un procés complex, delicat i excepcional. No tots els éssers vius que moren es transformen en fòssils, s'han de donar unes condicions molt específiques i tot i així no es segur que es pugui formar un fòssil. Les condicions necessàries són:
- Un soterrament ràpid que preserva les restes evitant l'acció d'éssers vius descomponedors i de l'ambient oxidant que afavoriria la ràpida destrucció dels teixits. És a dir, es fa necessari una atmosfera reductora lliure d'oxigen o anoxidant.
- Un sediment de gra fi que ompli totes les cavitats, esquerdes i detalls que pugui presentar l'ésser viu, d'aquesta manera la impressió final seria molt detallada. En aquest sentit el millor sediment és sempre el llim o l'argila.
- Una circulació contínua però lenta d'aigua saturada en sals minerals. Una alta concentració en sals minerals permet que sigui més fàcil la substitució dels components químics originals, més peribles per altres més duradors com la calcita o la sílice.
- Una situació d'estabilitat que es mantingui el temps suficient per afavorir la mineralització.
- I en general, que l'ésser posseeixi parts dures, prèviament ja mineralitzades pel propi organisme, ja que és més fàcil que fossilitzi l'esquelet d'un ésser viu que no pas els seus òrgans que normalment desapareixen fruit de l'acció de carronyers i descomponedors, encara que en casos excepcionals s'han pogut conservar com un miratge en la roca.
Veiem amb detall un exemple de fossilització:
En aquest exemple hem pogut veure com es forma un motlle intern i un motlle extern d'un ammonit, un cefalòpode que va aparèixer fa 400Ma i que es va extingir fa 65MA. Tot i així aquest no és l'únic procés de mineralització possible ja que podem distingir diverses possibilitats tot i que al cap i a la fi es redueix a una sola cosa: la substitució de la composició química de l'ésser viu per una altre:
- Precipitació de minerals dissolts en espais buits que ja existeixen, és el cas del re-ompliment dels forats que presenten les conquilles dels mol·luscs, això dóna lloc als anomenats motlles interns i ens donen informació de l'estructura fisiològica de l'ésser viu.
- Precipitació de minerals dissolts en espais buits deixats per matèria orgànica en descomposició. Quan les restes orgàniques d'un ésser viu com les seves parts orgàniques o els seus excrements queden soterrats, normalment amb el temps es descomponen deixant un forat que les aigües subterrànies poden omplir. Si aquesta aigua està prou saturada en sals minerals, aquestes sals precipitarien en el forat preservant la forma externa, l'empremta externa de les parts orgàniques ja descompostes. També es podria donar el cas que la mostra original contingués parts dures però que amb el temps han estat dissoltes, deixant una cavitat buida on precipiten altres minerals. És el que s'anomena un motlle extern de l'ésser viu.
- Re-ompliment de cavitats amb sediment. En aquest procés no implica tant la mineralització com els propis processos de sedimentació i diagènesi. La idea és que l'activitat d'un ésser viu com les seves petjades, els caus on vivien o els espais on construïen els seus nius poden ser omplerts per sediment que després litificaria.


- Substitució dels minerals pels quals està composta la mostra per uns altres diferents. A vegades, la closca, l'esquelet, o fins hi tot la matèria orgànica (és el cas de la fusta) pot ser modificada químicament fins al punt que el procés de fossilització substitueix la composició química original per una altre, per exemple les closques de carbonat càlcic poden ser substituïdes per sulfur de ferro (Pirita) o òxid de silici (sílice). I com es pot donar aquest procés? Bé, es requereixen unes condicions concretes de temperatura i composició química de les aigües subterrànies que afavoreixi la dissolució del mineral original i a la vegada faci precipitar l'altre mineral. Aquests fòssils solen ser molt apreciats per la seva bellesa. Dintre d'aquesta categoria també trobem els casos en els que la
mineralització ha afectat als teixits tous, de manera que les cèl·lules
s'han impregnat de minerals substituint literalment, la matèria orgànica
per matèria inorgànica. Aquesta mineralització es pot donar amb calcita
o sílice i és la que permet que trobem fusta perfectament fossilitzada
on es poden distingir totes les seves parts originals.

- Canvis en l'estructura cristal·lina dels minerals que la formen per uns altres més estables. Aquest procés es dóna quan les parts dures de la mostra original estan mineralitzades en substàncies amorfes d'una composició química concreta, com l'òpal o el nacre i aquesta, per processos de diagènesi canvia per altres estructures cristal·lines com la calcita, l'aragonita o la sílice. Així mateix també es pot donar el cas que la mostra original estigui feta d'aragonita i que aquesta canvii a calcita.
Bé, vist tot això arribem a la conclusió que la fossilització és un procés poc comú, difícil que succeeixi i delicat. Això ens fa pensar que tot i que el registre fòssil és ampli, es coneixen prop de 300000 espècies diferents aquesta deu ser una petita part de totes les espècies que han habitat el nostre planeta des que va sorgir la vida. Algunes estimacions parlen que hi ha existit més de mil milions d'espècies diferents de les quals només s'ha conservat un 0,000003% del total. És a dir, el registre fòssil és escàs i ens aporta una informació esbiaixada de com ha canviat la vida i el nostre planeta al llarg del temps.
I quina informació ens donen els fòssils?
Bé ens aporten informació climàtica de les condicions ambientals que predominaven mentre l'ésser estava viu,
condicions paleoclimàtiques. Ens aporten informacions
paleoestratigràfiques, ja que ens informen dels ambients de sedimentació, també ens donen informacions
biològiques, és a dir, en quines condicions vivien aquests animals, de que s'alimentaven, com vivien, quina era la seva fisiologia, etc. i sobre tot ens donen informacions
cronològiques, ja que ens ajuden a determinar l'edat de les roques i la seva cronologia. Es en aquest sentit en el que distingim el que s'anomenen
fòssils guia o
fòssils directors:
Un fòssil guia és el fòssil d'un organisme que va viure en un
període de temps curt, ja que evolucionava ràpidament i que a la vegada tenia una
amplia distribució per tot el món de manera que se'l pot trobar en moltes unitats estratigràfiques amb facilitat, és a dir, és també un fòssil molt
comú i abundant.
Aquest tipus de fòssil és molt apreciat en geocronologia ja que ens serveix per correlacionar estrats de diferents zones i ens aporten informació de l'edat de l'estrat i de les condicions ambientals i climàtiques en les que es va dipositar.
Va ser l'estudi dels fòssils guia el que va permetre assentar les bases de la teoria de la tectònica de plaques i a la vegada el que ens ha permès als geòlegs, junt amb la datació absoluta i la relativa a construir la taula dels temps geològics per poder portar un registre de tota la història del nostre planeta.
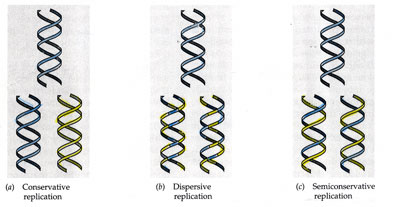

 Primer actua l'Helicasa que separa la doble hèlix trencant els ponts d'hidrogen, s'inicia la forca de replicació (es formen els replicons o bombolles de replicació).
Primer actua l'Helicasa que separa la doble hèlix trencant els ponts d'hidrogen, s'inicia la forca de replicació (es formen els replicons o bombolles de replicació).