Durant tota la segona unitat, en la que varem parlar de com s'estructurava la Terra, us vau estar preguntat com els geòlegs coneixien totes aquestes dades sobre l'interior del nostre planeta. Així mateix, ara que hem començat a parlar de la seva història, també us entren dubtes sobre com som capaços de saber com es va formar el planeta o com sabem la seva edat. Doncs bé, els geòlegs utilitzen una sèrie de mètodes que s'agrupen en dos tipus: els
mètodes directes i els
mètodes indirectes.
Mètodes directes
Quan parlem de mètodes directes ens referim a totes aquelles proves que podem efectuar
in situ, és a dir, tot allò que podem estudiar ja sigui en un laboratori o en un aflorament i que ens aporten resultats completament fiables i repetibles.
I quins són aquests mètodes? En principi us els podeu imaginar fàcilment:
- Anàlisi directe d'una roca recollida en un aflorament o en un sondeig, a través del qual podem saber la seva edat absoluta (per radiometria), la seva composició química, el contingut fòssil (si en té), el seu origen (si és orgànica, química o detrítica), etc.
- Estudi d'una mina, gràcies al qual podem conèixer en profunditat com es distribueixen els estrats, com varia la temperatura, com varia la pressió, quines són les condicions físiques que imperen a certes profunditats, etc.
- Estudi geològic d'una regió (mapa geològic), amb el qual podem conèixer aspectes relacionats amb l'ambient en el que s'han anat dipositant els diferents estrats, els processos tectònics que s'han donat, els paràmetres de forces que han actuat, la variació al llarg del temps dels ambients de sedimentació, etc.
A través de tots aquests mètodes podem conèixer una gran quantitat de dades del passat del nostre planeta, i en casos molt concrets, s'han pogut analitzar petits fragments del mantell litosfèric que han quedat atrapats en les vores convergents, a més d'altres roques abocades a l'exterior a través dels processos geològics interns i que es creu que provenen de zones molt profundes. Tot i així aquest coneixement directe queda molt delimitat a la superfície del nostre planeta. Per tant, la major part del volum del planeta és encara un misteri al qual només podem accedir a través dels anomenats mètodes indirectes.
Mètodes indirectes
Aquest grup reuneix tots aquells anàlisis que ens ofereixen indicis sobre l'estructura interna del planeta i per tant del seu origen i la seva formació.
De mètodes indirectes n'hi ha molts, però els més importants són els següents:
Els meteorits que cauen a la Terra ens donen molta informació de quina pot ser la composició del mantell i del nucli terrestre. La major part dels meteorits que cauen sobre el nostre planeta provenen del cinturó d'asteroides situat entre Mart i Júpiter. Es creu que aquest cinturó es va formar per causa de la destrucció d'un planeta que devia estar situat en aquella òrbita i segurament es va destruir per l'efecte contraposat de la gravetat del Sol i de Júpiter. El fet de saber que tots els planetes del sistema solar provenen del mateix núvol de roca i gas que va formar la Terra, ens fa pensar que tots els planetes rocosos haurien de tenir una composició similar. Al llarg del temps s'han analitzat gran quantitat de meteorits que es poden classificar en quatre grups diferenciats per la seva composició:
Siderits: composats principalment de ferro (Fe) i níquel (Ni). Són meteorits d'alta densitat. No s'ha trobat a la superfície de la Terra cap aflorament que indiqui que es puguin formar roques d'aquest tipus a l'escorça de forma natural. Si sumem aquesta informació amb l'alta densitat que presenten i sabent que durant la formació de la Terra els materials més pesants s'havien d'enfonsar i concentrar-se en el nucli, la presència d'aquests meteorits només té una explicació possible: són fragments d'un nucli planetari. Així doncs, l'estudi d'aquests siderits ens ajuda a saber com és el nucli terrestre.
Siderolits: Aquests meteorits estan composats per silicats metàl·lics i metalls. Per la seva estructura cristal·lina i la seva composició és creu que ens donen indicis de la composició de zones molt profundes del mantell inferior.
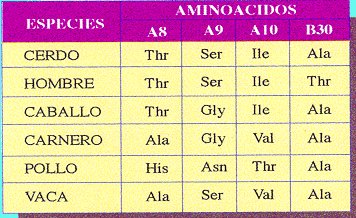
Les
condrites són meteorits no metàl·lics amb una composició molt similar a la de l'escorça terrestre. Estan composats principalment per silicats. S'han efectuat datacions radiomètriques de molts d'aquests meteorits que han donat com a resultats edats pels volts dels 4500Ma. Aquestes mesures corroboren l'edat aproximada de la formació del nostre planeta, ja que el més probable és que tant el nostre planeta com els asteroides dels quals provenen aquestes condrites es formessin al mateix temps. D'altra banda, les condrites contenen aminoàcids i altres molècules orgàniques. Aquest fet aporta solidesa a la teoria de la
panspèrmia com a possible origen de la vida en el nostre planeta.
Aquest mètode es basa en la mesura de la gravetat en diferents punts de la superfície de la terra buscant zones d'anomalies, ja sigui per augment de la densitat o per descens. Les mesures de les variacions gravimètriques han donat validesa a la teoria de la isostàsia que explica que l'escorça està formada per roques de baixa densitat que floten en equilibri sobre un fluid d'alta densitat.
Les anomalies positives fan pensar que hi ha més massa de matèria de la que hauria d'haver i les negatives que passa just el contrari. Si observem el mapa adjunt, podem veure com les zones d'anomalies positives solen coincidir amb relleus importants mentre que les zones de gravimetria negativa responen a conques on l'escorça està especialment aprimada.
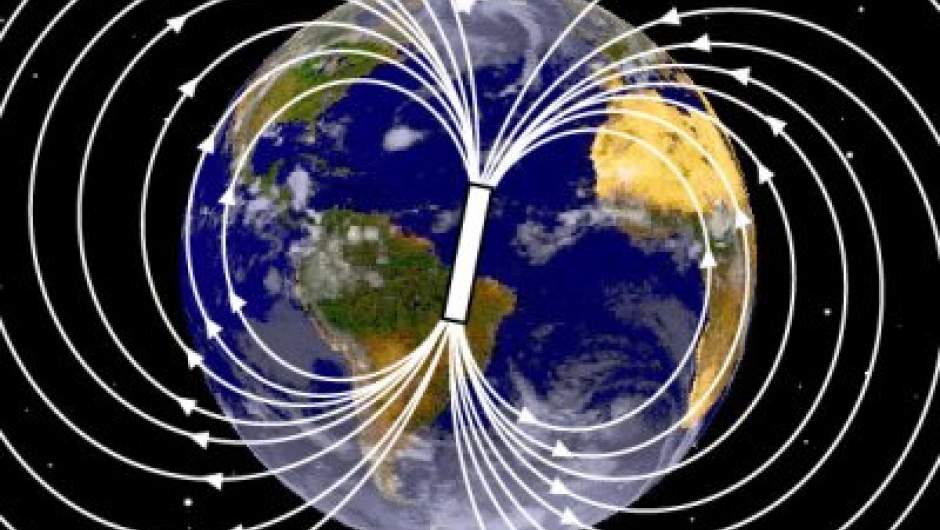
L'estudi de les anomalies magnètiques va permetre validar la teoria de l'expansió oceànica de la que ja vam parlar en el post sobre la
deriva continental. Així mateix, i a nivell local, les anomalies magnètiques ens ajuden a detectar roques susceptibles al magnetisme que poden tenir d'una banda interès econòmic i d'una altre interès científic.
Es basen en les propietats elèctriques que poden presentar algunes roques, ja sigui per la seva conductivitat i la resistència al pas del corrent elèctric, si són poroses i contenen aigua seran bons conductors i si hi ha anomalies elèctriques que pot indicar la presència de minerals d'interès econòmic.
Aquest mètode que es basa en la transmissió de les ones sísmiques generades desprès d'un terratrèmols a través de l'interior del nostre planeta, és el mètode més important per tal de conèixer com és el nostre planeta.
Es basa en la idea de que si la Terra fos uniforme, les ones sísmiques es transmetrien de la mateixa manera en totes direccions tal i com podem veure en la imatge següent:
A mesura que augmentava el coneixement sobre les ones sísmiques i augmentava la precisió dels sismògrafs els geòlegs es van adonar que hi havia tot una sèrie de zones fosques per on no es transmetien les ones sísmiques. L'única explicació que van trobar és que el planeta havia de tenir una configuració tal com aquesta:
En la imatge podem veure com es transmeten les ones sísmiques després d'un terratrèmol.
Normalment aquesta transmissió ve condicionada per la composició química del material que travessen i sobre tot i més important encara, la seva estructura física. Els sismògraf enregistren generalment dos tipus d'ones sísmiques: les
ones P i les
ones S.
Les
ones P són ones que vibren en la mateixa direcció en la que es mouen i es transmeten molt bé en materials rígids, sòlids, tot i que també es transmeten en materials fluids, ja que aquests també poden vibrar d'aquesta manera.
Les
ones S són ones que vibren verticalment a la direcció en la que es desplacen. De manera que si bé es desplacen molt bé a través de materials rígids, perden ràpidament intensitat en materials fluids fins al punt de desaparèixer completament.

Alguns aspectes que els geòlegs van tenir en compte a l'hora d'interpretar els resultats són que les ones P viatgen sempre més ràpidament que les ones S, que a més profunditat i més pressió més ràpidament es desplacen les ones sísmiques i que quan una ona, ja sigui P o S, passa d'un material amb determinades condicions físiques a un altre amb altres condicions aquestes és refracten de la mateixa manera que ho fa un raig de llum quan passa de l'aire a l'aigua.
Analitzant els resultats dels sismògrafs i tots els aspectes relacionats amb les ones sísmiques els geòlegs vam poder determinar quines eren les capes de la terra que ja vam estudiar en la unitat anterior:
Bé, fins aquí el post sobre els mètodes d'estudi. Ens hem deixat d'explicar la radiometria, que tot i formar part dels mètodes directes, ens serveix per explicar la datació absoluta que formarà part del pròxim post.